

What Is A Major Advantage Of Aplysia For Studies On The Physiology Of Learning?
Chapter 7: Learning and Memory
The analysis of the anatomical and physical bases of learning and memory is 1 of the great successes of mod neuroscience. Thirty years ago niggling was known almost how memory works, but now nosotros know a smashing bargain. This Affiliate volition discuss 4 bug that are central to learning and retention. Start, what are the different types of memory? Second, where in the encephalon is retentiveness located? One possibility is that homo memory is like to the memory scrap in a personal computer (PC), which stores all the memory in ane location. A 2nd possibility is that our memories are distributed and stored in different regions of the brain. Third, how does memory piece of work? What types of changes occur in the nervous system when a retention is formed and stored, are at that place particular genes and proteins that are involved in retention, and how tin can a memory terminal for a lifetime? 4th, is the issue of importance to many people, especially as we age: How can memory be maintained and improved, and how can information technology be stock-still when it is cleaved?
7.1 Types of Retentiveness
Psychologists and neuroscientists have divided memory systems into two wide categories, declarative and nondeclarative (Figure 7.one). The declarative retention system is the system of memory that is perhaps the about familiar. Information technology is the retentiveness arrangement that has a conscious component and it includes the memories of facts and events. A fact like 'Paris is the capital of France', or an event like a prior vacation to Paris. Nondeclarative memory, also called implicit retentivity, includes the types of memory systems that do non accept a conscious component merely are nevertheless extremely important. They include the memories for skills and habits (eastward.m., riding a bicycle, driving a car, playing golf game or tennis or a piano), a phenomenon called priming, uncomplicated forms of associative learning [east.g., classical conditioning (Pavlovian conditioning)], and finally simple forms of nonassociative learning such as habituation and sensitization. Sensitization will be discussed in detail afterwards in the Chapter. Declarative retentiveness is "knowing what" and nondeclarative retention is "knowing how".
| Figure seven.one |
7.2 Testing Memory
| Figure 7.2 |
| Figure 7.3 |
Everyone is interested in knowing how well they remember so let usa take a simple memory test. The examination (Figure 7.2) will nowadays a list of xv words, so there will be a interruption and you will be asked whether yous remember some of those words. Sorry, y'all have to put your pen downward for this exam and exercise not read further in the Affiliate until you consummate the exam.
This retentiveness test chosen the DRM test subsequently its creators James Deese, Henry Roediger and Kathleen McDermott. It was not meant to exist a trick, but to illustrate a very interesting and important feature about memory. We like to call up that retentivity is similar to taking a photograph and placing that photograph into a filing cabinet drawer to be withdrawn later (recalled) as the "retention" exactly the way it was placed there originally (stored). But memory is more than like taking a flick and violent it up into pocket-sized pieces and putting the pieces in different drawers. The memory is then recalled by reconstructing the memory from the individual fragments of the memory. The reason so many individuals incorrectly believe that "sweetness" was on the list is considering there were so many other words on the listing that had a sweet connotation. "Declining" this exam is actually not a bad consequence. Individuals with Alzheimer's disease more often than not do not say that "sweetness" was on the list. They cannot make the normal associations involved in the recall of a memory.
The word list gives insights into memory processing and retrieval, just information technology is non a really good examination of "raw" retentiveness ability because information technology can be afflicted by distortions and biases. To avoid these problems, psychologists take developed other memory tests. One is the object recognition test (Effigy 7.3) to examination declarative retentivity. This examination is also skillful because, as we volition see later, it can even be used on animals. The test involves presenting a subject with two different objects and they are asked to retrieve those objects. At that place is a pause and then ii objects are shown over again, one of which is new and the other having been shown previously. Subjects are asked to identify the novel object, and to exercise so, they need to recollect which ane was shown previously. A somewhat related test is the object location test in which subjects are asked to recall the location of an object on a two-dimensional surface.
Examples of nondeclarative memory, such as associative learning, can be tested by pairing one stimulus with another and later testing whether a subject field has learned to make the association betwixt the two stimuli. The classical instance is the image developed by the Russian physiologist Ivan Pavlov, which is now called classical or Pavlovian conditioning. In classical conditioning (Figure 7.4), a novel or weak stimulus (conditioned stimulus, CS) like a audio is paired with a stimulus like food that by and large elicits a reflexive response (unconditioned response, UR; unconditioned stimulus, United states) such equally salivation. After sufficient grooming with contingent CS-Us presentations (which may be a single trial), the CS is capable of eliciting a response (conditioned response, CR), which often resembles the UR (or some attribute of it).
| Effigy 7.4 |
vii.iii Localization of Memory
Now permit us plow to this event nigh where is memory located. There are iii basic approaches.
- Imaging. Mod imaging techniques like fMRI (functional magnetic resonance imaging) or PET (positron emission tomography) allows one to "see" areas of the encephalon that are agile during specific brain tasks. If a subject is placed in an fMRI scanner and given a memory test, one can make up one's mind what areas of the brain are active, and that activity presumably is related to where in the brain the memory is processed and/or stored.
Figure 7.5
PET brain scan during an object location exam. (from A. Thousand. Owen, et al., J. Cog. Neurosci. 8:half-dozen, 588-602, 1996.)Figure seven.5 illustrates an example of a PET scan of an individual who is performing an object location examination. The color code is such that the brighter, redder regions bespeak increased brain activity. The almost active region is the hippocampus. In discussions of retentivity, the hippocampus is mentioned repeatedly because it is a major part of the brain involved in declarative memory function. This illustration conspicuously indicates that the hippocampus is involved in object location memory. But equally we will see soon, it is non where all memories are stored.
- Encephalon lesions. In this experimental procedure, small parts of the brains of mice or rats are surgically removed or chemically inactivated and the animals are systematically examined to determine whether the lesion afflicted whatsoever memory organisation.
- Brain illness and injury. Here scientists take advantage of individuals who have had unfortunate brain injuries, for example, through stroke or through a brain tumor in a specific surface area of the brain. If one finds a memory arrears in the patient, it is probable that the region of the brain that was injured is involved in that memory.
A classic written report on localization of retentiveness was the upshot of surgery performed on Henry Molaison, a patient who was but known to the scientific community as "H.M." until his death in 2008. H. K. is famous in neuroscience literature because his brain provided major insights into the localization of memory function. In the 1950's, H.M. was diagnosed with intractable epilepsy, and while at that place are pharmacologic treatments, in some cases the just treatment is to remove the portion of the brain that is causing the seizures. Consequently, H.Thou.'southward hippocampus was removed bilaterally. Figure seven.half dozen (right) is an MRI of a normal individual showing the hippocampal region, whereas Figure 7.half dozen (left) shows a MRI of patient H.Yard. subsequently the removal of the hippocampus.
Figure 7.6
Bran scans of H.M. (left), and a normal individual (right). (Copyright © 1997 past Suzanne Corkin, used with permission of The Wylie Agency LLC.)
Before the operation, H.M. had a fine memory, but after the operation, H.K. had a very severe memory arrears. Specifically, afterwards the functioning H.Thou.'s ability to grade whatsoever new memories for facts and events was severely impaired; he had swell difficulty learning whatsoever new vocabulary words; he could not recollect what happened the day before. And then if H.One thousand. had an interview the 24-hour interval following a previous interview, he would accept little or no memory virtually the interview or events during information technology. This study clearly indicated that the hippocampus was disquisitional for memory germination. Only whereas H.M. had swell difficulty forming new memories for facts and events, he nevertheless had all of his old memories for facts and events. Specifically, he had all his childhood memories, and all of his memories prior to the operation. This type of memory deficit is chosen anterograde amnesia. (In contrast, retrograde amnesia refers to loss of old memories.) The studies on H.M. clearly indicated that whereas the hippocampus is critical for the formation of new memories, it is not where the old memories are stored. It is now known that those quondam memories are stored in other parts of the brain, such as in the frontal cortex. The process by which an initially labile memory is transformed into a more enduring form is chosen consolidation. This process involves the memory being stored in a dissimilar role of the brain than the initial site of its encoding.
H.M. was also interesting in that while his ability to form new memories for facts and events was severely dumb, he could form new memories for skills and habits. While he could course new memories for skills and habits, he did not know that he had the skills! He had no awareness of the retention; he couldn't declare that he had it. This finding clearly indicated that the memory for skills and habits are not formed in the hippocampus. Collectively, we learned from these studies on H.K. and other patients that retention is distributed throughout the nervous organization, and different brain regions are involved in mediating different types of memory.
Figure 7.7 summarizes many decades of research on the anatomical locus of memory systems. The medial temporal lobe and structures like the hippocampus are involved with memories for facts and events; the striatum is involved with memories for skills and habits; the neocortex is involved with priming; the amygdala is involved with emotional memories; and the cerebellum with simple forms of associative learning. Lower brain regions and the spinal string contain even simpler forms of learning. In summary, retention is not stored in a unmarried place in the brain. It is distributed in different parts of the brain.
| Figure 7.seven |
7.iv Mechanisms of Memory
Model systems to report retention mechanisms
| Figure seven.8 |
Much of what has been learned almost the neural and molecular mechanisms of learning and memory have come from the use of and then chosen "model systems" that are acquiescent to cellular analyses. 1 of those model systems is illustrated in Figure 7.8A.Aplysia californica is found in the tidal pools forth the coast of Southern California. It is most six inches long and weighs nigh 150 grams. At beginning glance it is an unpromising looking creature, but neuroscientists accept exploited the technical advantages of this beast to gain cardinal insights into the molecular mechanisms of memory. Indeed, the pioneering discoveries of Eric Kandel using this animate being were recognized by his receipt of the Nobel Prize in Physiology or Medicine in 2000. Aplysia have three technical advantages.
Showtime, it exhibits uncomplicated forms of nondeclarative (implicit) learning like classical (Pavlovian) conditioning, operant conditioning and sensitization.
2d, Aplysia have a very simple nervous arrangement. Compared to the 100'due south of billions of nerve cells in the human brain, the unabridged nervous organisation of this brute only has about 10,000 cells. Those cells are distributed in different ganglia similar the ane illustrated in Figure 7.8B. Each ganglia like this 1 has only near 2,000 cells, however it is capable of mediating or decision-making a number of different behaviors. This means that any 1 behavior tin be controlled by 100 neurons or fifty-fifty less. One has the potential of working out the consummate neural excursion underlying a behavior, and so, later on training the brute, the neural circuit can be examined to identify what has inverse in the excursion that underlies the memory.
Third, the ganglia contain neurons that are very big. Figure 7.8B shows a ganglion under a dissecting microscope. It is about 2mm in diameter. The spherical structures throughout the ganglia are the prison cell bodies of individual neurons. Each neuron is identifiable and has a unique localization and part. A related advantage is that individual neurons can be removed and placed in culture medium where they can survive for many days. Indeed, multiple neurons can exist removed from the ganglia and they reestablish their normal synaptic connections, thereby providing a very powerful experimental system to study the physiology of nerve cells and the properties of the connections betwixt them.Figure 7.8C shows an example of a sensory neuron (small cell to the right) and a motor neuron (large cell to the left) in culture. In the micrograph information technology is possible to see the shadow of a microelectrode that has impaled the sensory neuron, and the shadow of a microelectrode that has impaled a motor neuron for performing intracellular recordings.
Sensitization, a simple course of nondeclarative learning amenable to detailed cellular analyses
| Effigy 7.9 |
| A. | B. | C. |
| Figure 7.10 | ||
Figures 7.9 and vii.10 illustrate a uncomplicated beliefs exhibited by the animal and a simple course of learning called sensitization. The beast is tested past stimulating its tail with a weak electric daze (vii.ix) or a weak mechanical tap (7.10). These stimuli elicit defensive reflex withdrawals of the body, which includes the tail and nearby sites such as the gill and a fleshy spout called the siphon. In response to examination stimuli delivered every five minutes, the withdrawals are fairly reliable. They are about the same elapsing each fourth dimension (Figures 7.9B, C, 7.10A). But if a potent noxious stimulus (eastward.m., an electrical shock) is delivered to another part of the brute such as its trunk wall, subsequent test stimuli to the tail give enhanced responses (Figure seven.9B and 7.10B). This is an instance of a simple course of learning called sensitization. It is divers equally the enhancement of the response to a test stimulus as a upshot of delivering a strong generally noxious stimulus to the animal. In a sense, the fauna is learning that it is in a "fearful" environment. Sensitization is a ubiquitous form of learning that is exhibited by all animals including humans.
Neural excursion and mechanisms of sensitization
- Neural circuit. We tin take advantage of the large nerve cells of Aplysia, and the ability to make intracellular recordings from them, to piece of work out the underlying neural excursion. Figure 7.11 illustrates a simplified view of the key components of the underlying neural circuit. Stimulation of the skin activates sensory neurons (SN) (simply one of which is illustrated here) which brand glutamatergic excitatory synaptic connections (triangles) with motor neurons (MN). If the summated synaptic input to the motor neurons is sufficiently big, the motor neurons volition be activated and action potentials will propagate out of the ganglion to cause an eventual contraction of the muscle. So stimulation of the skin excites sensory neurons, the sensory neurons activate motor neurons, and motor neurons contract the muscles. Besides, it should be evident that the greater the activation of the motor neurons, the greater will be the subsequent reflex response. This reflex in Aplysia is similar to the knee jerk or stretch reflex mediated by similar circuitry in the vertebrate spinal cord.
Effigy 7.11
Neural circuit for the defensive withdrawal reflex.
- Mechanisms of sensitization. Sensitizing stimuli pb to the release of the neurotransmitter serotonin (5-HT) (represented past cell labeled IN and colored in regal on Figure 7.11). five-HT modulates the strength of the connection betwixt the sensory neuron and the motor neuron. An activity potential in the sensory neuron before the learning produces a small excitatory postsynaptic potential (EPSP) in the motor neuron (Effigy 7.12A). But, after delivering the sensitizing stimulus, an activity potential in the sensory neuron leads to a larger synaptic potential in the motor neuron (Figure 7.12C). A larger synaptic potential in the motor neuron increases the probability that the motor neuron will exist activated to a greater extent and produce a larger contraction of the muscle (i.eastward., sensitization).
I principle well-nigh learning and memory derived from studies of this simple animal, and this principle holds truthful in our brains as well, is that learning involves changes in the strength of synaptic connections betwixt neurons. Learning is not due to a reorganization of the nervous system or the growth of new neurons. What has changed is that the strength of a previously existing connection is modified.
Now we tin can take this analysis one step further and enquire what are the biochemical mechanisms that underlie learning and retention. We will split the discussion into ii temporal domains of memory; curt-term retentiveness and long-term memory. We take already discussed different types of retention such every bit declarative and nondeclarative retention. In that location are too different temporal domains of retention. Curt-term memories are similar the memory for a telephone number that last several minutes, and long-term retentivity are memories that final days, weeks or a lifetime.
Figure 7.12A
Earlier sensitization. Slide the blue brawl to control the animation.Figure 7.12B
During sensitization. Slide the blue brawl to control the blitheness.Figure 7.12C
After sensitization. Command the blitheness by sliding the blue ball.
- Mechanisms of short-term sensitization. The mechanisms for the brusk-term retentiveness for sensitization are illustrated in Figure seven.12B. The sensitizing stimulus leads to release of the neurotransmitter five-HT. 5-HT binds the 2 types of receptors on the sensory neuron; one is coupled to the DAG/PKC arrangement, and the other is coupled to the circadian AMP/PKA system. These are the same general cascades that you learned in biochemistry. Learning mechanisms accept evolved to co-opt some of the biochemical machinery that are already present in all cells used them specifically for a retentiveness mechanism in nerve cells. The protein kinases exert two types of actions. Showtime, they regulate the backdrop of unlike membrane channels (the pocket-size gates on the illustration (Effigy 7.12) correspond membrane channels that underlie the initiation and the repolarization of the activity potential). Consequently afterward a sensitizing stimulus, the corporeality of calcium that enters the synaptic final during an action potential and causes the release of transmitter will be enhanced. In addition, the modulation of the membrane channels leads to an increment in the excitability of the sensory neuron and as a result a greater number of action potentials will be elicited by a test stimulus to the pare. 2nd, the kinases regulate other cellular processes involved in transmitter release, such as the size of the pool of synaptic vesicles available for release in response to the influx of Ca2+ with each activity potential. Finally, 5-HT leads to changes in the properties of the postsynaptic motor neuron. Specifically, 5-HT leads to an increase in the number of glutamate receptors. The consequences of these processes can be seen by comparing the strength of the synaptic connection produced by a single action potential before (Effigy vii.12A) and afterwards (Figure vii.12C) sensitization. The specific details of all the currents and processes are not critical. However, information technology is of import to know the general principles. One principle is that learning involves the date of second messenger systems. Here both the poly peptide kinase C (PKC) and the protein kinase A (PKA) systems are involved. This is a fairly general principle. In every case of learning that has ever been examined, whether vertebrate or invertebrate, 2d messenger systems are engaged. A second principle is that memory involves the modulation of neuronal membrane channels. These can include channels that directly regulate transmitter release (i.e., Ca2+ channels in the presynaptic neuron), channels that regulate neuronal excitability, and channels that mediate synaptic responses in the postsynaptic neuron. A third principle is that cyclic AMP is one of the disquisitional 2nd messengers that is involved in memory. Given this information, yous tin begin to recall virtually how retentivity could exist improved based on your knowledge of the underlying biochemistry.
Nosotros have discussed a mechanism for a brusk-term retention. Information technology is "short-term" because the memory is transient and that is so considering the underlying biochemical changes are transient. The elapsing of the memory is dependent on how long the various substrate proteins (e.g., membrane channels) are phosphorylated. PKA will only exist activated for a short time later on a brief stimulus because cyclic AMP will be degraded and PKA levels will decrease. Protein phosphatases will remove the phosphate groups on the substrate proteins that are "storing" the memory.
Figure 7.thirteen
Structural changes in sensory neurons associated with long-term sensitization. (Modified from 1000. Wainwright et al., J. Neurosci. 22:4132-4141, 2002.)
- Mechanisms of long-term sensitization. There are two major differences between brusque-term and long-term memories. Long-term memories involve changes in protein synthesis and factor regulation, whereas brusk-term memories exercise not. And, long-term memories in many cases involve structural modifications. Figure seven.13 illustrates examples of the processes of 2 sensory neurons that have been filled with a dye, one from an untrained creature and one from a trained brute. Shown are the thick axonal procedure of the neuron and many fine branches. Along the branches are seen pocket-size dot-like swellings or varicosities. These varicosities are the presynaptic terminals of the sensory neurons that make contact with other neurons like the motor neurons. (The motor neurons cannot be seen because only the sensory neurons were filled with the dye.) Part B of Figure seven.13 shows an instance of a sensory neuron that has been injected with a dye in an untrained animal, and Part A shows one that has been filled with a dye 24 h after sensitization training. At that place is a major difference between these two neurons. The neuron from the trained animal has a greater number of branches and a greater number of synaptic varicosities than the neuron from the untrained animate being. Therefore, long-term memory involves changes in the structure of neurons including growth of new processes and synapses. So, to the extent that you think annihilation about this material on retentivity tomorrow, or next week, or next twelvemonth, it will be because structural changes in synapses are beginning in your brains!
Figure vii.fourteen
Genes implicated in long-term sensitization.Given that long term memory involves changes in gene expression, a major goal of neuroscientists is to place the specific genes and proteins that are involved in long-term memory. Figure vii.14 illustrates some of the genes and the proteins that are involved in long-term sensitization. Note that campsite, ane of the second messengers involved in the curt-term retentiveness, is also involved in the induction of long-term retention. But now, in addition to its effects on the phosphorylation of membrane channels, camp, through PKA, phosphorylates transcription factors such equally CREB (cAMP responsive element binding protein). Transcription factors like CREB, when phosphorylated, are capable of regulating gene expression, which leads to changes in the expression of proteins that are important for inducing and maintaining the long-term changes in synaptic strength and therefore the long term retention.
Note that at that place is not a single "magic retentivity gene" – rather, the consecration and maintenance of memory, fifty-fifty in a single neuron, involves the appointment of multiple genes and proteins that act synergistically to change the properties of the neurons and regulate the backdrop of the neuron and the force of the synapse. Too note that changes in gene expression do non occur all at once – there are different phases. Some changes in gene expression occur early on, some even 24 hours after the learning occurs.
Long-term potentiation (LTP): A probable synaptic machinery for declarative retention
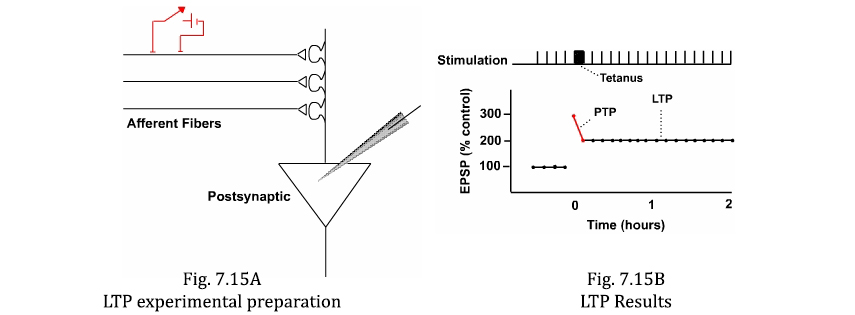
An enduring form of synaptic plasticity called long-term potentiation (LTP) is believed to be involved in many examples of declarative retentivity. It is present in the hippocampus, which is known to be involved in declarative memories. LTP can be studied in brain slice preparations where an electric shock (test stimulus) can exist delivered to afferent fibers and the resultant summated EPSP can be recorded in the postsynaptic neuron (Figure 7.15A). If the pathway is repeatedly stimulated (e.one thousand., every infinitesimal), the amplitude of EPSP is abiding (Figure 7.15B).
Delivering a brief ane-sec duration railroad train of high frequency (100 Hz) stimuli (i.e., the tetanus) to the afferent nerve produces two types of enhancement in the postsynaptic neuron. Offset, at that place is a transient facilitation called post-tetanic potentiation (PTP) that dies away after several minutes. Second, following the PTP is a very enduring enhancement of the EPSP chosen LTP. LTP is the kind of mechanism necessary to shop a long-term retentiveness (Effigy vii.15B).
Figure seven.sixteen
Animation of the induction and expression of LTP.The NMDA-type glutamate receptor is critical for some forms of LTP, in particular LTP at the CA3-CA1 synapse in the hippocampus. The postsynaptic spines of CA1 neurons have two types of glutamate receptors; NMDA-type glutamate receptors and AMPA-blazon glutamate receptors (Figures 7.16A). Both receptors are permeable to Na+ and K+, simply the NMDA-type has two additional features. Showtime, in addition to beingness permeable to Na+, it likewise has a significant permeability to Ca2+. Second, this channel is normally blocked by Mgtwo+.
Even if glutamate binds to the NMDA receptor and produces a conformational change, there is no efflux of K+ or influx of Na+ and Ca2+ because the channel is "plugged upward" or blocked by the Mg2+. Thus, a weak examination stimulus will not open this channel because information technology is blocked past Mg2+. A weak test stimulus volition produce an EPSP, but that EPSP will be mediated by the AMPA receptor. It is every bit if the NMDA receptor were non even there.
Now consider the consequences of delivering a tetanus (Figure 7.16B). During the tetanus, in that location will exist spatial and temporal summation of the EPSPs produced by the multiple afferent synapses on the common postsynaptic cell (Figure vii.15A). Consequently, the membrane potential of the postsynaptic neuron will be depolarized significantly, much more and then than the depolarization produced by a single afferent test stimulus. Because the inside of the prison cell becomes positive with the large synaptic input, the positively charged Mg2+ is repelled by the within positivity and is "thrust" out of the channel. At present the channel is unplugged and Ca2+ can enter the spine through the unblocked NMDA receptor. The Ca2+ that enters the cell activates various protein kinases, which then trigger long-term changes. One component of the long-term change is the insertion of new AMPA receptors into the postsynaptic membrane (Figure 7.16C). Therefore, after the tetanus, the transmitter released from the presynaptic neuron past a test stimulus volition bind to a greater number of receptors on the postsynaptic neuron. If more than receptors are jump and hence opened, a larger (potentiated) EPSP (i.e., LTP) volition be produced (Effigy seven.16C). In addition to an increase in the number of postsynaptic AMPA receptors, in that location is prove that a greater amount of transmitter is released from the presynaptic neurons. The combination of the presynaptic and postsynaptic effects would act synergistically to increase the size of the synaptic potential in the postsynaptic neuron. Note that this case of a synaptic mechanism for declarative retentivity bears some similarity to the synaptic mechanism for the example of nondeclarative memory (sensitization) discussed previously. Although the specific details differ, both involve activation of second messenger systems and regulation of membrane channels. Therefore, at a fundamental mechanistic level, there does not appear to exist meaning differences betwixt the 2 major classes of memory systems. The major difference appears to be the brain region and the neural circuit and into which the learning mechanism is embedded.
7.5 Enhancing Retentivity
Figure 7.17
A data plot of enhanced memory in transgenic mice.With a knowledge of some of the genes and proteins involved in memory, nosotros can use this information to try to both test the role of specific proteins in memory and also to improve retentiveness. One experimental style of approaching the upshot is to use transgenic technology in which a gene of interest can be over expressed in an animal by introducing it into an egg cell. When the offspring develop into adults, their performance on memory tests can be examined. An example of this approach is illustrated in Figure vii.17. Here the office of the NMDA receptor was examined by Joe Tsien and his colleagues, who were then at Princeton University. If NMDA receptors are important for the induction of LTP, and LTP is important for declarative memory, i would expect that animals that had a greater number of NMDA receptors would learn more readily. NMDA receptors were over expressed in mice and the mice were tested on the object discrimination examination that was discussed earlier in the Chapter.
To assess the functioning of a mouse on the object recognition task, the experimenter measures the amount of fourth dimension for some predefined period the mouse spends exploring the i object, versus the amount of time the mouse spends exploring the other object. If the mouse remembers that it had seen one of the objects previously, it will spend more time exploring the novel one. As illustrated in Effigy 7.17, i hour afterwards the initial presentation of the objects, the mice exercise very well on the examination. Indeed, they are correct about 100% of the time. They know the novel object. Nonetheless, 1 twenty-four hours later the retentivity operation is rather poor, and later on three days it is fifty-fifty worse. By i week, mice show no recognition memory.
What about the mice that received the extra NMDA receptors? Now one day after grooming they accept perfect retention! And so the extra receptors have led to an improved memory performance. That'southward the good news – but the bad news is that the retention is no improve ane week later. This somewhat disappointing finding should not be surprising. Although NMDA receptors are important in retentivity, they are not the whole story. Equally indicated earlier in the Chapter, retentiveness involves the synergistic appointment of multiple genes and proteins. So to improve retention further, it will be necessary to manipulate multiple genes. At the present time it is hard to do and then, just, it probably will go possible in the near future. It volition also be possible to over limited genes of interest in targeted areas of the human being brain. The future for treating individuals with retentiveness disabilities looks very promising.
This blitheness by Graduate students Julia Loma and Natalia Rozas De O'Laughlin of the Neuroscience Graduate Programme at McGovern Medical School at UTHealth explains the concept of synaptic plasticity. It placed third in the 2011 Countdown Order for Neuroscience Encephalon Sensation Video Contest. Test Your Knowledge
- Question 1
- A
- B
- C
- D
- E
A 50-year old patient with recent damage to the hippocampus from a stroke would probable accept all of the following deficits EXCEPT:
A. Difficulty learning new facts
B. Difficulty describing a recent effect
C. Difficulty learning a new vocabulary word
D. Difficulty recalling a babyhood memory
Eastward. Difficulty remembering a face up
A 50-year old patient with contempo harm to the hippocampus from a stroke would likely have all of the following deficits EXCEPT:
A. Difficulty learning new facts This reply is Incorrect.
The hippocampus is involved in declarative memory including the memory for facts.
B. Difficulty describing a recent event
C. Difficulty learning a new vocabulary word
D. Difficulty recalling a childhood retention
E. Difficulty remembering a face
A 50-year old patient with recent impairment to the hippocampus from a stroke would likely have all of the post-obit deficits EXCEPT:
A. Difficulty learning new facts
B. Difficulty describing a recent outcome This respond is INCORRECT.
The hippocampus is involved in declarative memory including the memory for recent events.
C. Difficulty learning a new vocabulary discussion
D. Difficulty recalling a childhood retentiveness
Due east. Difficulty remembering a face
A 50-twelvemonth old patient with recent damage to the hippocampus from a stroke would likely have all of the following deficits EXCEPT:
A. Difficulty learning new facts
B. Difficulty describing a recent event
C. Difficulty learning a new vocabulary discussion This answer is INCORRECT.
The hippocampus is involved in declarative memory including the memory for vocabulary words (semantic memory).
D. Difficulty recalling a childhood memory
Due east. Difficulty remembering a face
A 50-year old patient with contempo damage to the hippocampus from a stroke would likely have all of the post-obit deficits EXCEPT:
A. Difficulty learning new facts
B. Difficulty describing a recent event
C. Difficulty learning a new vocabulary word
D. Difficulty recalling a childhood retentiveness This reply is CORRECT!
The hippocampus is involved in the formation of new memories, but not in the storage of one-time memories after they have been consolidated.
E. Difficulty remembering a confront
A fifty-year former patient with recent damage to the hippocampus from a stroke would probable accept all of the following deficits EXCEPT:
A. Difficulty learning new facts
B. Difficulty describing a contempo event
C. Difficulty learning a new vocabulary word
D. Difficulty recalling a childhood memory
E. Difficulty remembering a face This answer is INCORRECT.
The hippocampus is involved in object recognition.
- Question 2
- A
- B
- C
- D
Short term memories can involve all of the following processes EXCEPT:
A. Regulation of gene expression
B. Activation of second-messenger systems
C. Modulation of membrane channels
D. Modulation of transmitter release
Curt term memories can involve all of the following processes EXCEPT:
A. Regulation of factor expression This reply is Right!
Regulation of gene expression is associated with long-term memories and not short-term memories.
B. Activation of second-messenger systems
C. Modulation of membrane channels
D. Modulation of transmitter release
Short term memories can involve all of the following processes EXCEPT:
A. Regulation of cistron expression
B. Activation of 2nd-messenger systems This respond is Wrong.
Activation of 2d-messenger systems such as cAMP is associated with brusque-term retentivity.
C. Modulation of membrane channels
D. Modulation of transmitter release
Short term memories tin involve all of the following processes EXCEPT:
A. Regulation of gene expression
B. Activation of second-messenger systems
C. Modulation of membrane channels This reply is INCORRECT.
Both voltage-gated and transmitter-gated channels are associated with short-term memory.
D. Modulation of transmitter release
Short term memories can involve all of the following processes EXCEPT:
A. Regulation of factor expression
B. Activation of 2d-messenger systems
C. Modulation of membrane channels
D. Modulation of transmitter release This answer is INCORRECT.
Changes in synaptic strength are associated with curt-term memory.
- Question three
- A
- B
- C
- D
- E
Classical conditioning is an example of:
A. Semantic memory
B. Episodic memory
C. Implicit memory
D. Declarative memory
East. Nonassociative memory
Classical conditioning is an instance of:
A. Semantic memory This answer is INCORRECT.
Semantic memory is a type of declarative memory, whereas classical conditioning is a type of nondeclarative (implicit) retention.
B. Episodic retentivity
C. Implicit retentiveness
D. Declarative memory
E. Nonassociative retention
Classical conditioning is an example of:
A. Semantic retentivity
B. Episodic memory This answer is INCORRECT.
Episodic retention is a type of declarative retention, whereas classical workout is a blazon of nondeclarative (implicit) retention.
C. Implicit memory
D. Declarative memory
Eastward. Nonassociative memory
Classical conditioning is an example of:
A. Semantic memory
B. Episodic memory
C. Implicit memory This respond is CORRECT!
D. Declarative memory
East. Nonassociative retentivity
Classical workout is an instance of:
A. Semantic memory
B. Episodic memory
C. Implicit memory
D. Declarative memory This answer is Incorrect.
Classical conditioning is an example of nondeclarative retentivity.
E. Nonassociative memory
Classical conditioning is an example of:
A. Semantic memory
B. Episodic memory
C. Implicit memory
D. Declarative retentiveness
East. Nonassociative memory This answer is Wrong.
Classical workout is a form of associative learning, which is in contrast to examples of nonassociative retentiveness like sensitization.

Donations to Neuroscience Online will help fund development of new features and content.

Source: https://nba.uth.tmc.edu/neuroscience/m/s4/chapter07.html
Posted by: bergergaceaddly.blogspot.com
0 Response to "What Is A Major Advantage Of Aplysia For Studies On The Physiology Of Learning?"
Post a Comment